大元山植物生態之研究
── 章樂民 ──
資料來源:林業試驗所報告第070號
五、植物社會之研究(Study in Plant Communities)
植物社會乃一複雜有限之有機體,具有一定之發育過程與組織,受環境因子所控制,乃環境因子支配下之產物,植物社會最大單位為植物群系(Plant Formation),同一群系主耍以氣候因子與植物生活形(Life Forms)(指優勢種與次優勢種而言)皆田類似下而區分為若干群叢(Plant Association),群叢之下因土壤關係又分為若干植物帶(Zonation)即喬木下分為灌木,草本,著生植物,苔蘚,地衣等,此種縱向之劃分構成多層次(Layer)之樹冠,交互出現,第一層樹冠之疏密,可決定第二、三層之植被種類,頗為重耍,植物社會之區分各有不同之分類形式,大元山之植物社會依作者意見,應稱為常綠木本植物社會,雖然各林帶氣候土壤及植物種類略有差異,但其植物生活形與形態上之構造則大致相若,其區分如下:

1.亞熱帶降雨林群系(Subtropical rain forest formation)
本島熱帶降雨林,僅限於本島南部之恆春半島,即楓港溪,阿塱衛溪以南之地區,四季暖熱,年雨量平均在3,000mm以上,乾燥期甚短,植物生育旺盛,代表此種降雨林主要植物除龍腦香科(Dipterocarpaceae)無分佈外,主耍者以棕梠科(Plamae),大戟科(Euphorbiaceae),桑科(Moraceae)蕁麻科(Urticaceae)等植物,其他地區熱帶降雨林之發育甚為貧弱,所見者均應屬於亞熱帶降雨林,其分佈僅限於海拔高500m以下,除常見之熱帶植物外,並參與若干常綠闊葉樹林帶之樟科(Lauraceae)及殼斗科植物(Fagaceae),實際上可稱為常綠闊葉樹林延長林帶,故其分佈範圍甚狹,本省低海拔地常由於人口稠密,開墾,火災,伐採等現象,甚為普遍,亞熱帶降雨林已難保持其真正熱帶雨林之植物景色。茲就植物群叢(Associatfon),組成層次(layers),胸高直徑與株數分配,天然更新(Reproduction),生態演進(Suecession)等分述如下:
(I)植物群叢概述(Simple description of plant association)
構成大元山亞熱帶降雨林之主要植物草叢(Plant Association)分述如下:
山黃麻─榕中途群叢(Trema orientalis-FicusSpp., Associes)分佈於低海拔100~700m,土壤略乾燥,寫照充足之開墾地或伐採跡地,為第二期森林之早期發 生之植物,在山腳部占最多數,廣汎分佈於全島。
白匏仔中途單叢(Mallotuspaniculatus,Consocies)
分佈於海拔高100~8,000m,土壤乾燥或濕潤均適於生長,平地山麓以至於中海拔之開墾地或伐採跡地佔多數,當其他中性闊葉樹侵入後成被壓木時,則逐漸淘汰,為第二期森林最習見之陽性植物。
桫欏群叢(Cyathea-taiwaniana-Alsophila pustulosa, Association)
分佈於海拔高300~800m,北向蔭濕環境中之常綠闊葉樹林下部,成大面積散生狀態,尤其溪流峽谷最為繁茂,為亞熱降雨林特有之木本羊齒,普遍分佈於全島。
木苧蔴中途團集(Boehmeria densiflora, Colony)
分佈於海拔高100~500m,向陽乾燥環境,溪傍河邊崩壤地或伐採跡地,屢屢成群生狀態,普遍分佈於全島。
山香圓─山龍眼群叢(Turpiniaformosana-Heliciaformosana, Association)
分佈海拔高300~700m,濕潤闊葉樹林中,為亞熱帶降雨林第二層樹冠,一般高度均在4m以下,為亞熱帶降雨林主要宭叢之一。
長尾尖錐栗─黃杞群叢(Castanopsis longicaudata-Engelhardtiaformosana,
Association)
分佈於海拔高300~700m,土壤深厚而潤濕環境中,構成亞熱帶降雨林第一層樹冠,亦為主要群叢之,中性植物社會,為氣候極盛相(climatic climax)
芭西木群叢(Barthea formosana, Association)
分佈於海拔高500~700m,土壤稍濕潤,亞熱帶降雨林鬱閉略破壞,陽光折射林內之林下灌木,散生或成團集狀態,其分佈範圍不若常綠闊葉樹林內之普遍。
火炭母藤中途團集(Polyonum chinense, Colony)
分佈於海拔高300~500m,向陽略乾燥環境中,或鬱閉破壞後林下略有發現,尤其林綠路傍最多,成塊狀分佈。
菅草中途群叢(Miscanthus spp. Associes)
分佈於海拔高400~1,800m,土壤乾燥貧瘠環境中,開墾地,火燒跡地,崩壞地最多,林緣林隙亦有發生,屢屢成大面積分佈,為全省林地習見群叢之一。
芒箕骨中途群叢(Dicranopteris linearis, Consocies)
分佈於海拔高300~500m,向陽乾燥土壤貧瘠環境中局部出現,多生於酸性之紅土,路傍林緣特多。
(2)組成曆次(Structure layers)
構成大元山亞熱帶降雨林組成層次,可分為喬木層(Trees layer),灌木層(Shrubs layer),草本層(Herb layer),及著生植物苔蘚,地衣等,18個樣區測定結果,茲列各表以示之。

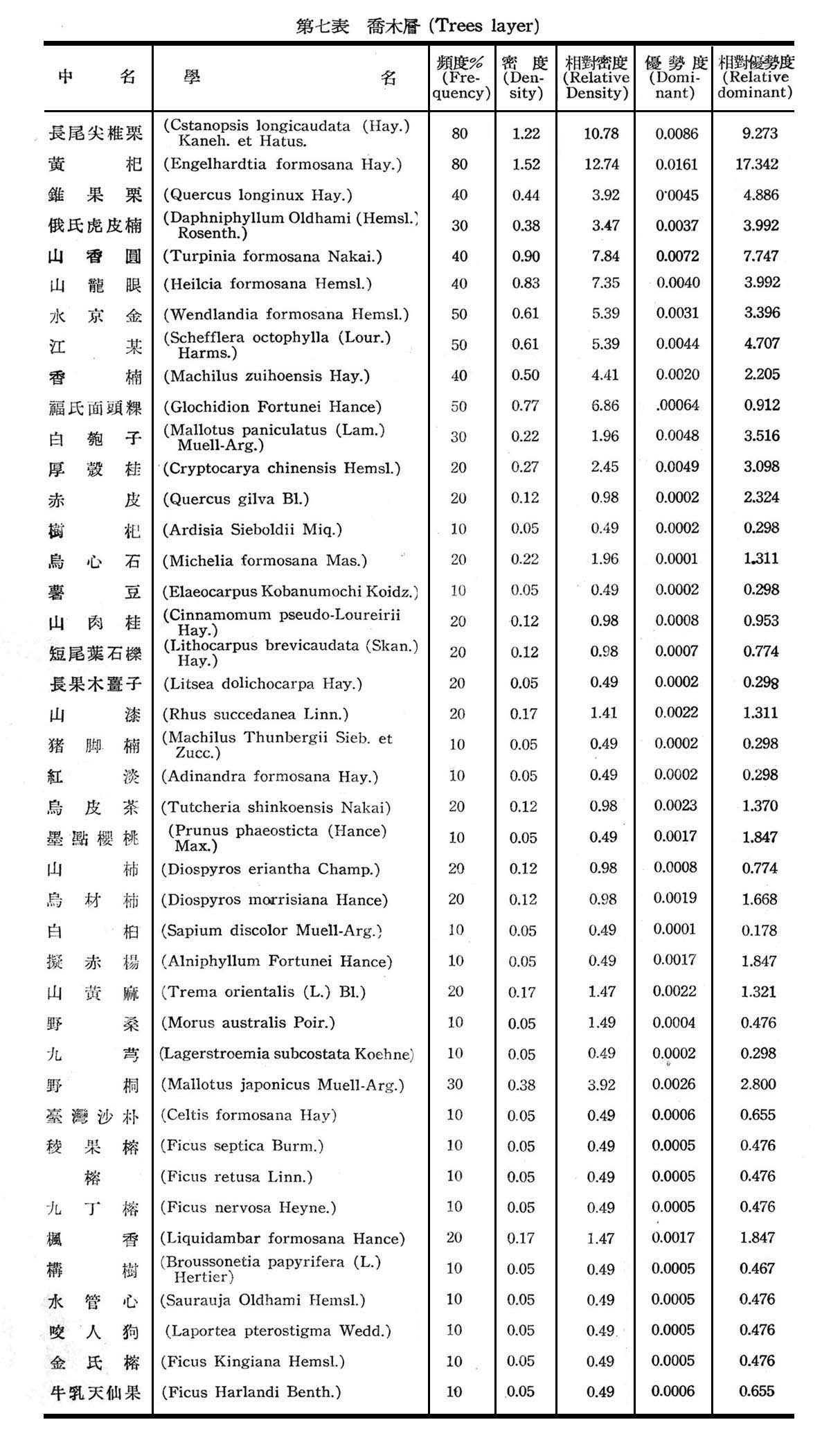


觀上列第七─九表構成亞熱帶降雨林計喬木層42種,灌木層31種,草本層21種,著生植物(羊齒植物)16種,苔蘚植物一種,地衣一種,由表7所示頻度,密度,優勢度均以長尾尖錐栗及黃杞為最大,為優勢種(Dominant Species),錐果櫟,江某,俄氏虎皮楠,山龍眼,山香圓,水京金,厚殼桂,福氏面頭粿,白匏仔等次之,為次優勢種(Co-dominant Species),故構成大元山亞熱帶降雨林,仍以長尾尖錐栗─黃杞群叢為主體。
(3)胸高直徑與株數分配:
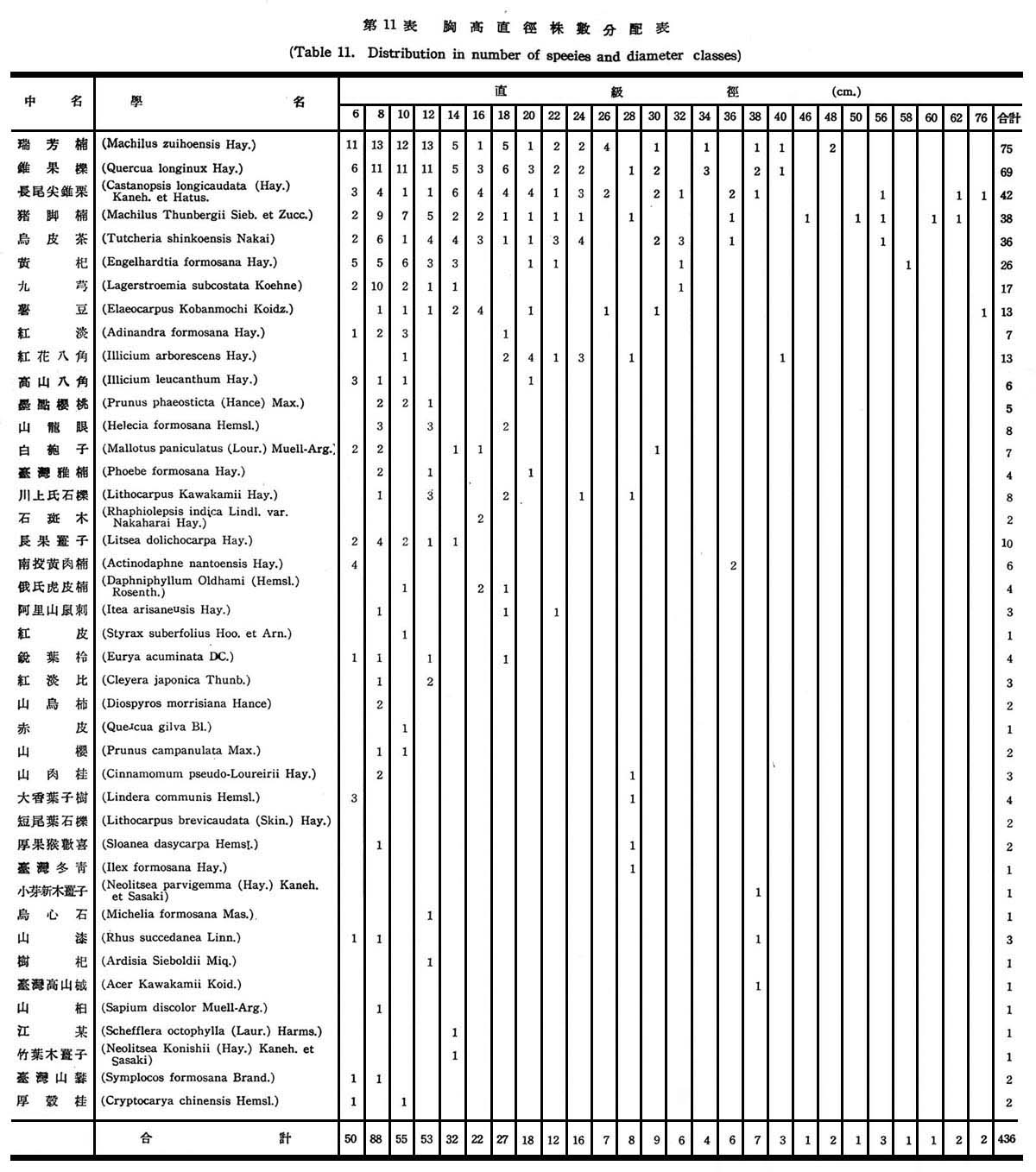
根據18個樣區測定結果,在目前狀態下亞熱帶降雨林之胸高直徑與株數分配如下圖所示:
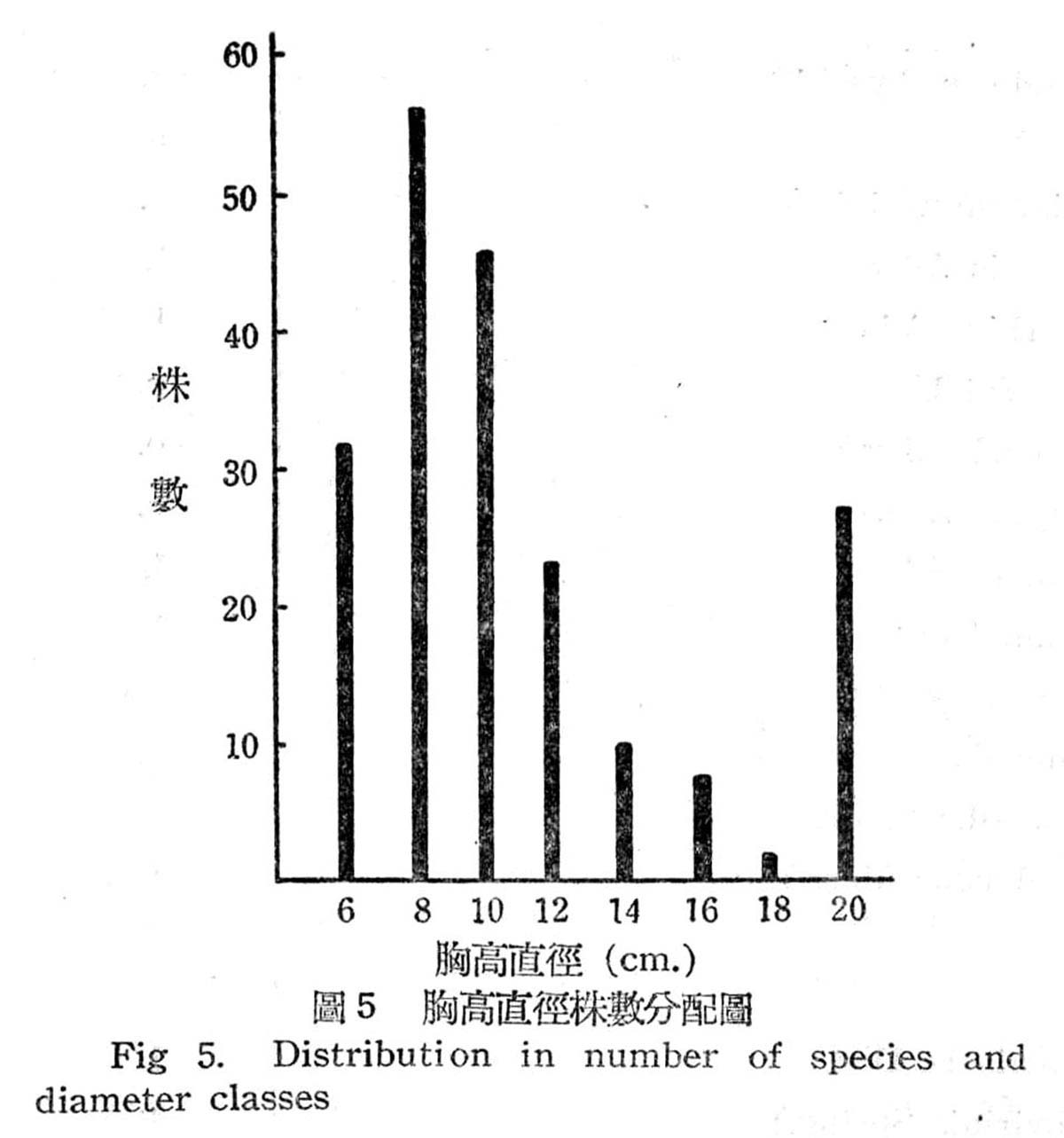
由圖5顯示此乃擇伐後之林型,蓋本省低海拔森林受人為破壞甚為普遍,直徑15cm以下佔77%,15~20cm佔23%,最大直徑未超過20cm以上者,故幼齡木佔絕大多數,壯齡木佔極少數,此森林早期已遭破壞,目前森林不過為伐採後之第二期幼年林。
(4)天然更新概況:(Natural Regeneration)
18f固樣區以測定其天然更新概況,以確定是否已達極盛點(climax.),結果如下表所示。
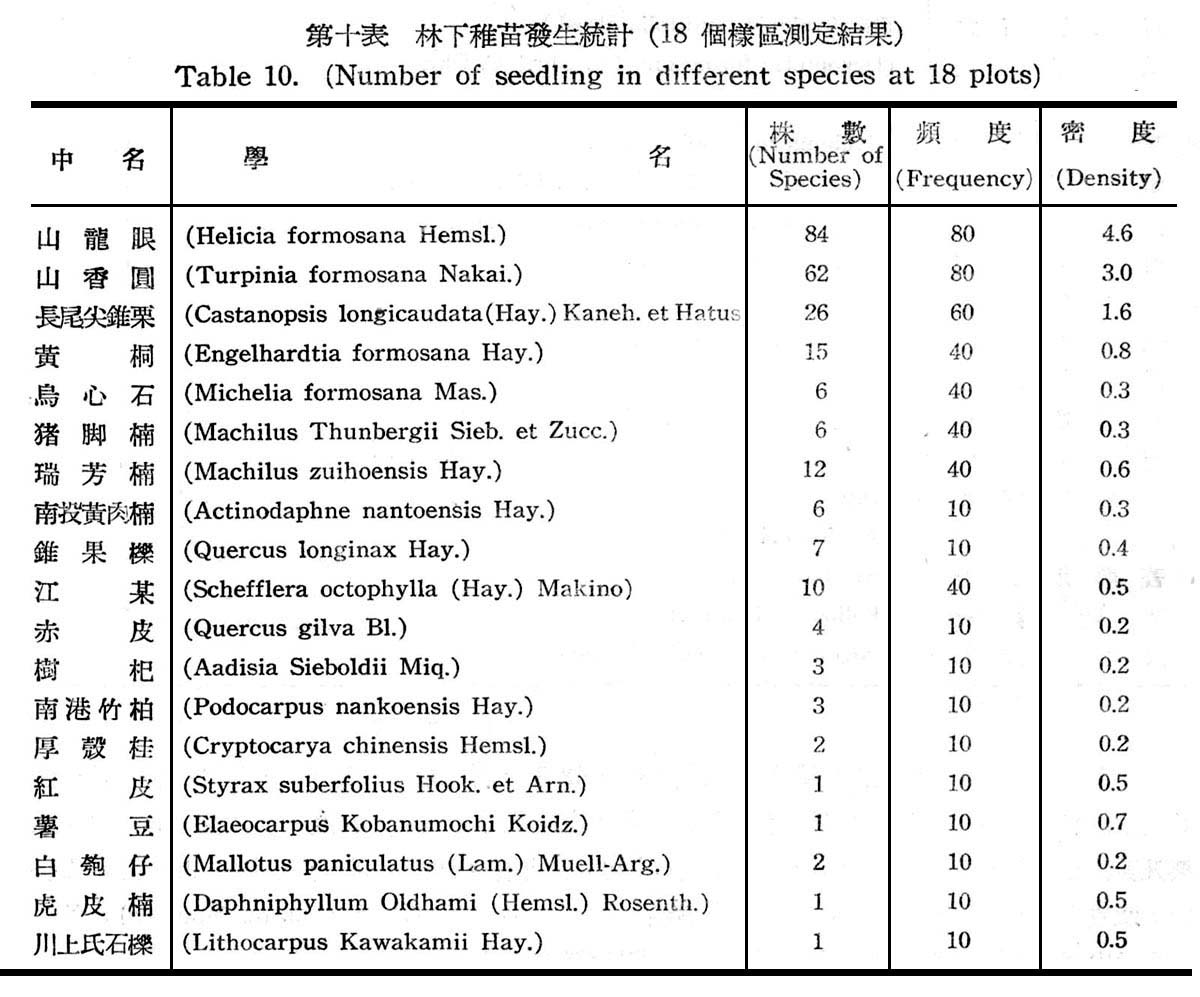
觀上表可知林下稚苗發生之頻度及密度,均以山龍眼及山香圓佔最大,其次則為長尾尖錐栗及黃杞,查山龍眼及山香圓均為亞熱帶降雨林習見之植物,近於半陽性者,但仍繼績延續而佔優勢,反之長尾錐栗及黃杞為以後侵入種近於中性植物,在第一層樹冠雖佔優勢,但其次代幼苗稚樹發生並不佔優勢,而較山龍眼及山香圓反居於劣勢,顯示長尾尖錐栗及黃杞,尚未達極盛植物社會(climax),此可確定仍在演變過程中,唯長尾尖錐栗及黃杞,將仍不失為該處之極盛植物社會主要份子。
(5)生態演進之推論(Plant Succession)
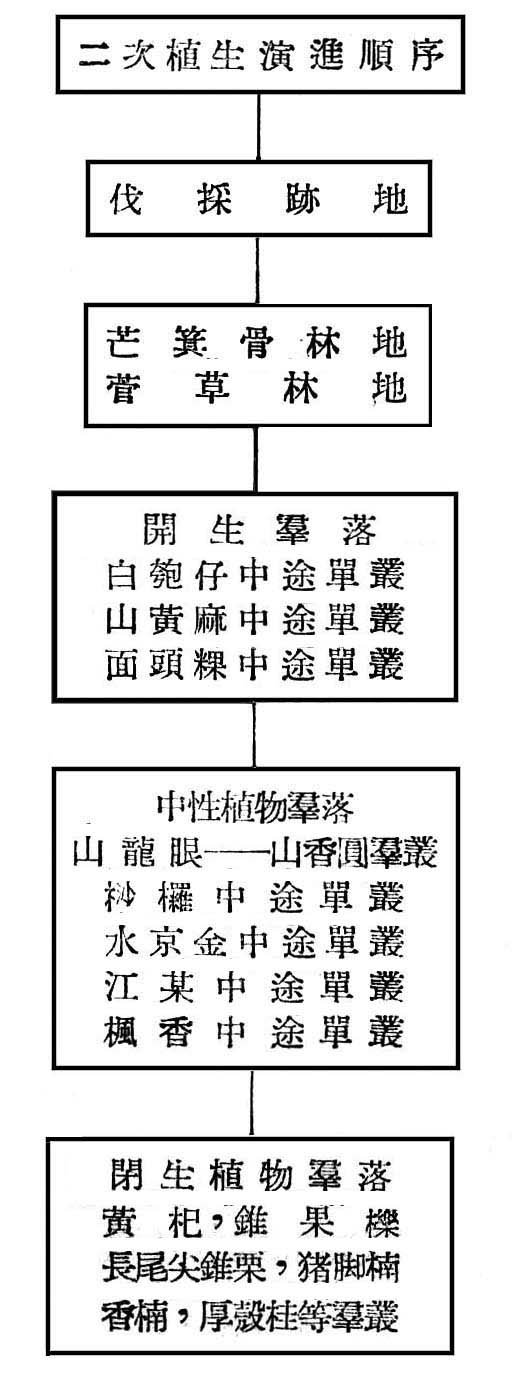
基於上列之分析,則大元山亞熱帶降雨林之生態演進,可作如是之推論,即大元山海拔高300~700m,亞熱帶降雨林早期受人為破壞,目前之第二期森林為伐採後發生之幼齡林,二次植生正在劇變(Secondary Succession),誘導二次植生演進之因子,不外乎濫伐,火災,開墾等因子所引起。原始植物演進Primary Succession)不外乎乾性植生列(Xerosere)與楊性植生列(Hydrosere),此地區植物生態之演進,應屬於乾性植生列殆無問題,且其演變過程中必須有一定之各時期(succession stage),且須較長久時間,至二次植物演進不須經一定時期與時間,當一已有森林之林地由於人為破壞與伐採後,林地暴露,土壤乾燥而固結,首先發生者為芒箕骨(Dieranopteris Spp.)及菅草(Miscantllus Spp.)植物群落所覆蓋,此等植物維持時間並不長久,即為開生群落(Open Community)如白匏仔(Mallotus Spp.),山黃麻(Trema Sp.),面頭粿(Glochidion Spp.)等極端陽性植物所侵入,此等陽性植物發育互相競爭結果,樹冠交叉而蔽蔭,反而造成不適於本身生存之環境,同時空氣中水分之下降,土壤由乾燥而濕潤,加以枯枝落葉之腐爛以增加土壤腐植質,立地略有改善,而介於中性與陽性植物間如山龍眼,山香圓,桫欏(Cyatllea-Alsophila Spp.),水京金,江某等相繼侵入,生長競爭結果,使陽陸植物逐漸淘汰,樹冠交叉更蔽蔭,土壤更為濕潤,日照減少,腐植質堆積,立地環境大為改善,則閉生群落(Close Community)如黃杞,錐果櫟,長尾尖錐栗,豬腳楠,香楠等次第侵入乃形成目前森林,在不受人為干涉情況下,目前森林將維持一較長久時期,唯根據現狀觀察知早期陽性植物雖逐漸減少,而介於陽性植物與中性植物間之山龍眼,山香圓,水京金等依然佔優勢,尤其次代之幼苗及稚樹頻度密度均佔最大,故此森林尚未達極盛點(Climax)。茲列一簡表以示其演變順序:
2.常綠闊葉樹林群系(Everngreen broad-leaved forest formation)
常綠闊葉樹林為本島主要植物群落型之一,樹種最為豐富,其分佈範圍恆在2500m以下,其中並有小部份為落葉喬木林外,主要以樟科(Lauraceae),殼斗科(Fagaceae),為主體,並參與若干山茶科(Theaceae),及山礬科(Symplocaceae)植物,在經濟價值上言似較亞熱帶降雨林高,大元山海拔高100~700m,列為亞熱帶降雨林已如上述,海拔高700~1300m則應列為常綠闊葉樹林帶,茲分述如下:
(1)群叢概述(Simple description of plant association)
瑞芳楠─錐果櫟群叢(Machilus zuihoensis-Quercus longinux, Association)
分佈於海拔高700~1300m,土壤濕潤肥沃山腹部佔最多,為構成大元山常綠闊葉樹林之主體,佔領面積廣潤,近於極盛植物社會。
長尾尖錐灤―豬腳楠群叢(Castanopsis longicaudata-Machilus Thunbergii, Association)
分佈於海拔高700~1300m,亦為構成大元山常綠闊葉之主體,土壤潤濕肥沃山腹佔最多,分佈面積亦廣,近於極盛極物社會。
南投黃肉楠一烏心石群叢(Actinodaphne nantoensis-Michelia formosana, Assocaiation)
分佈於海拔高700~1,300m溪谷向蔭濕環境中,土壤肥沃地多小團集狀態分佈,唯佔領範圍稍狹。
九芎群叢(Largerstromea subcostata, Association)
分佈於悔拔800~1,000m,近山腳溪傍甚為蔭濕之森林中,往往成小團集狀態,故其分佈範圍甚狹。
赤楊一管草中途群叢(Alnus formosana-Miscanthus sinensis, Associes)
分佈於海拔高700~1,000m,向陽而土壤略乾燥,火災後林地,崩壞地,溪傍或河床段丘地,多有發生,尤其崩壞地往往成大面積植物社會。
杜鵑―厚皮香群叢(Rhododendron Spp.-Ternstromia gymnantllera, Association)
分佈於山稜線部份,海拔高1,800~2,000m,陽光強烈,基岩暴露,土壤乾燥而貧瘠形成高山灌木林狀態,分佈範圍甚狹。
臺灣擦木─鹽膚木中途群叢(Sassafras Ramdaiense-Rhus javanica var. Roxburghii,Associes)
分佈於海拔1,200~1,500m,土壤乾燥陽光充足,開墾地或崩壞地,林綠部份往往成小團集狀態,分佈面積狹。
臺灣擦木一山胡椒中途群叢(Sassafras randaiense-Litsea cubeba, Associes)
分佈於海拔高1,000~1,300m,向陽乾燥土壤,往往成小團集狀態,以路傍開墾地,伐採跡地,露出地多有發現,分佈面積狹。
虎刺中途單叢(Damnacanthus formosanus, Consocies)
分佈於海拔高700~1,300m常綠闊葉樹林鬱閉咯破壞,而陽光折射林內略乾燥環境中,最習見之林下小灌木,多成小面積塊狀分佈,但範圍甚廣。
芭西木群叢(Barthea formosana, Association)
分佈於海拔高700~1,300m,亦為常綠闊葉樹林鬱閉良好之林下小灌木,但有時往往與虎刺混生成塊狀分佈,範圍亦廣。
玉山箭竹單叢(Pleisblastus niitakyamensis, Consociation)
分佈於海拔高1,000~1,300m,日照充足土壤乾燥環境中,伐採跡地,無立木地,往往大面積分佈。
菅草中途群叢(Miscanthus Spp., Associes)
分佈於海拔高400~1,800m,日照充足土壤乾燥貧瘠環境,崩壞地,開墾地,火燒地,溪傍河床地,均有大面積分佈。
塗氏苔團集(Carex Dunni, Clan)
分佈於海拔700~1,300m,常綠闊葉樹林鬱閉良好,土壤濕潤肥沃,最常見之林下地被植物,往往成小群落狀態,分佈範圍甚廣。
冷水花群叢(Pellionia scabra, Association)
冷水花─冷清草群叢(Pellionia Spp-Pilea Spp., Association)
分佈於海拔高700~1,300m,為常綠闊葉樹林林下主要地被植物,鬱閉良好,土壤深厚肥沃林地最為常見,有時成單叢,有時與冷水花(Pellinoia seabra),三萼冷水花(Pellinia trilobulata)阿里山冷清草(Piled brevicornuta)能高冷清草(Pilea nokozanensis)混合成群落分佈。
臺灣秋海棠單叢(Begonia laciniata var. formosana Consociation)
海芋單叢(Alocasia macrorrhiza, Consociation)
闊葉赤車使者群叢(Elatostema edule Association)
分佈於海拔高700~1,300m,常綠闊葉樹林林下習見之地被植物,蔽蔭土壤濕潤環境中成群生狀態。
紫苧麻團集(Villebrunea fruticosa, Colony)
分佈於溪傍,林緣,向陽而濕潤土壤成團集狀態,但分佈面積小。
(2)組成層次(Structure layers)
構成大元山常綠闊葉樹林組成層次30個樣區測定結果如下列各表所示。
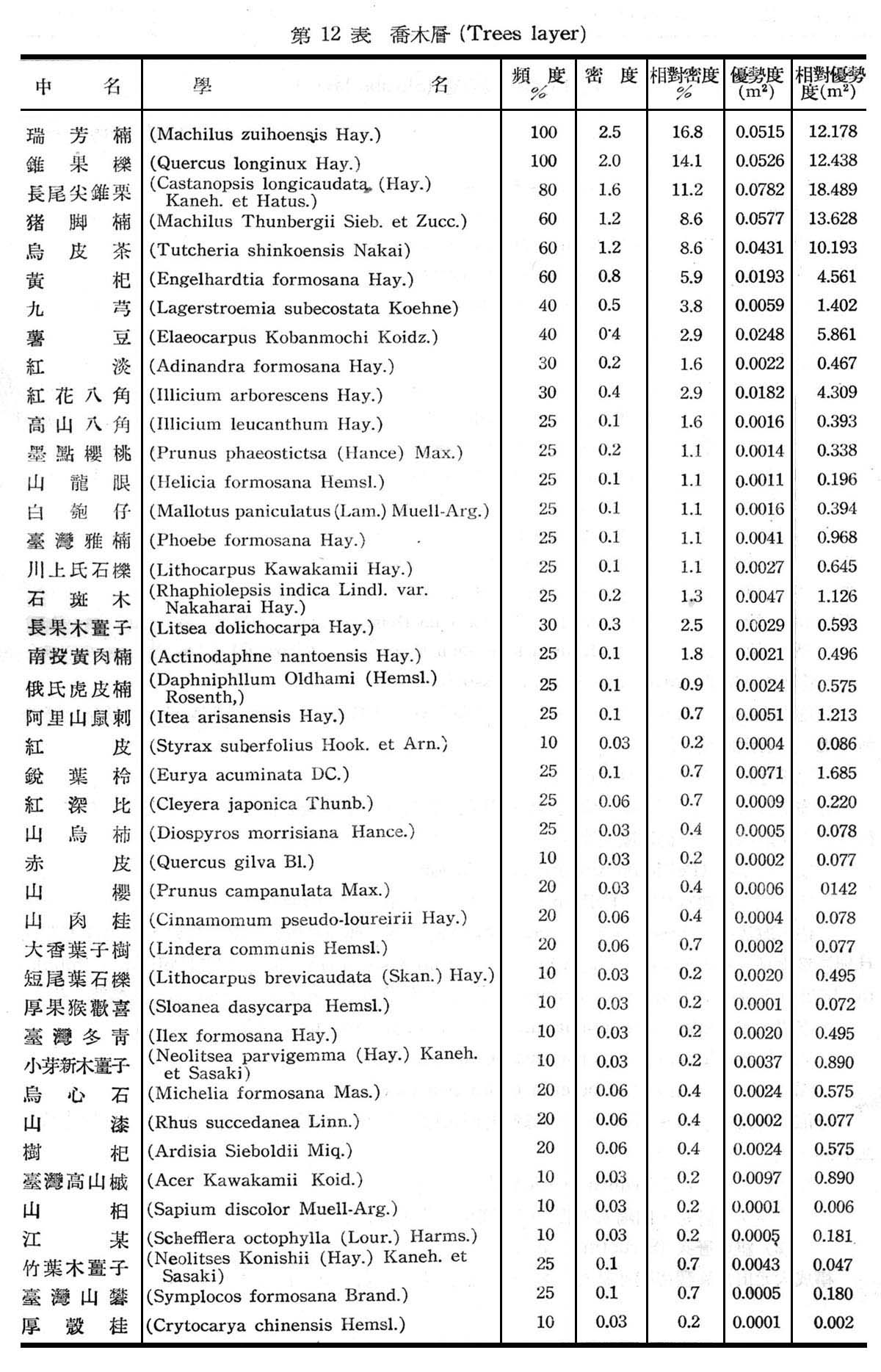


由上列12~14表統計可知構成大元山常綠闊葉樹料組成層次喬木層計42種,灌木層計36種,草本層計24種,羊齒植物計23種,苔蘚植物計5種,由第12表所示頻度,密度,優勢度等均以香楠,錐果櫟,長尾尖錐栗豬腳楠,烏皮茶佔優勢為優勢種(DominantsSpecies),黃杞,九芎,薯豆為次優勢種(Co-dominants Species),香楠,錐果櫟,長尾尖錐栗,豬肉楠為構成常綠闊葉樹林之主要群叢。
(3)胸高直徑與株數分配:
根據30個樣區測定結果,常綠闊葉樹林在目前狀態下,其胸高直徑與株數分配如第6圖所示。
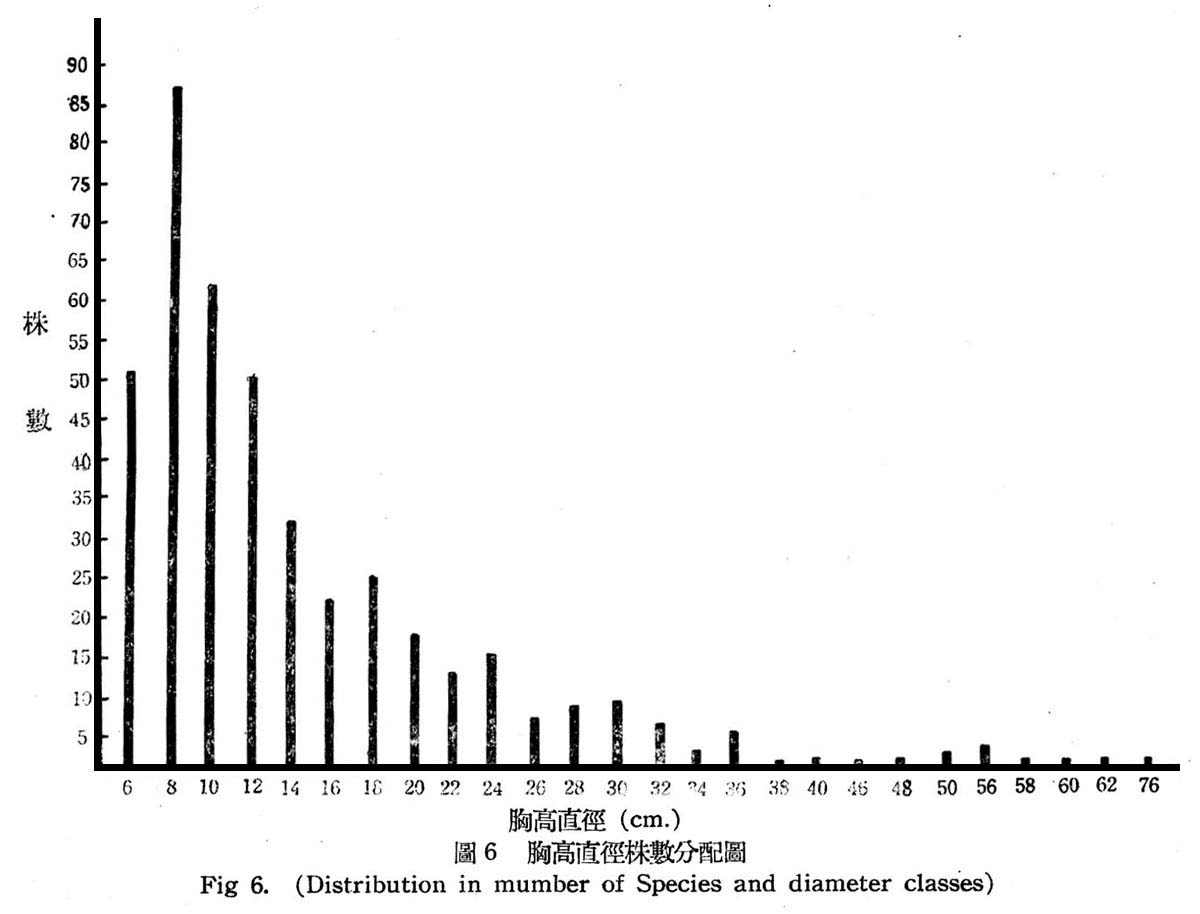
由圖6所示,大元山常綠闊葉樹林與下部亞熱帶降雨林之林型,大致相同,仍為早期受人為破壞與伐採,依目前現狀觀察仍第二期發生之幼齡林,胸高直徑12cm以下者占57.5%,12~38cm者占40%,40cm以上者占2.5%,即幼齡木占大多數,壯齡木占少數,老齡木幾已伐採殆億,此顯森林早期巳遭破壞。
(4)天然更新(Natural Regeneration)概況:
30個樣區以測定常綠闊葉樹林在天然情況下,其天然更新稚樹幼苗發生概況,以確定是否已達極盛點(Climax),結果如下表所示。
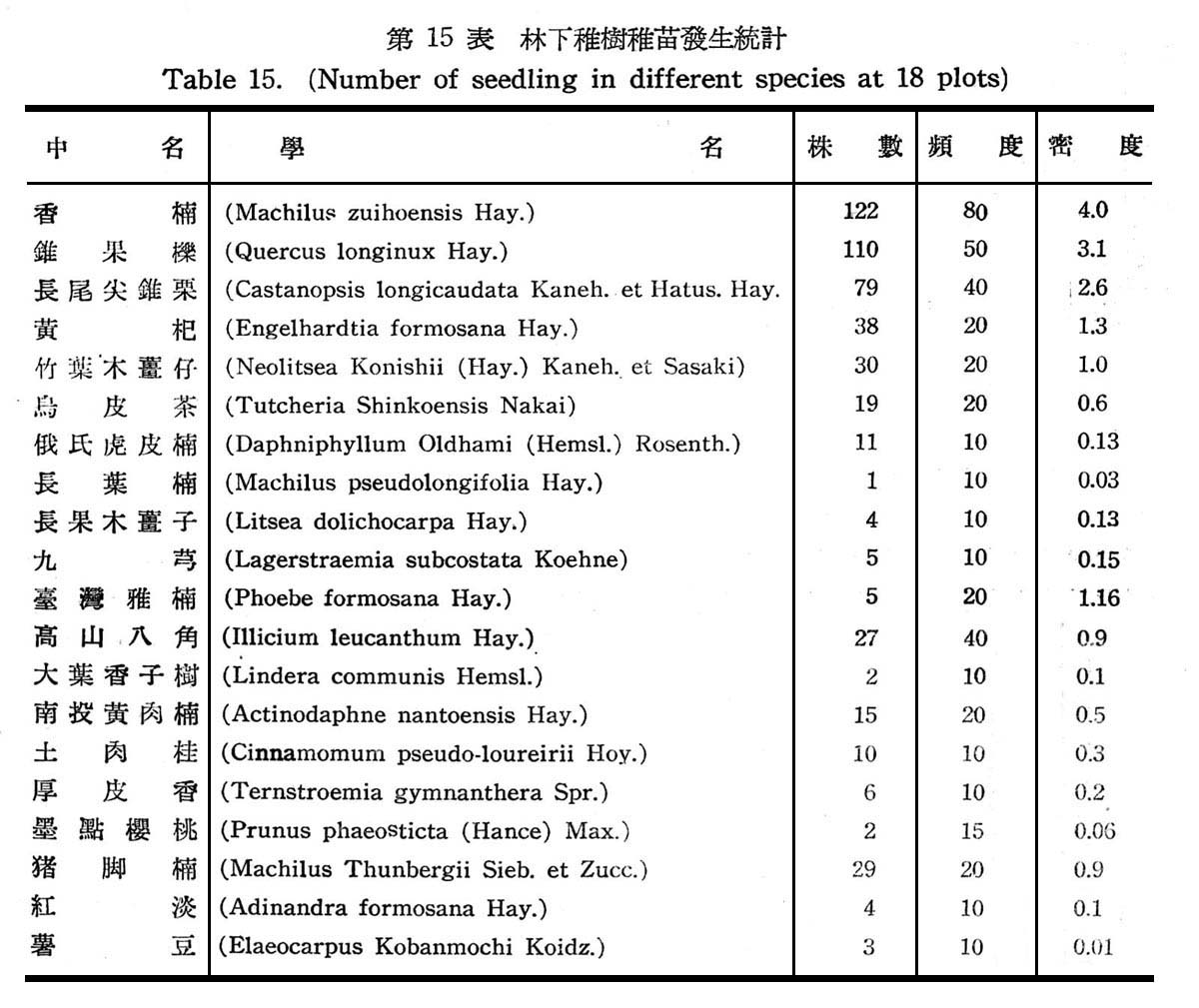
觀上表統計,香楠及錐果櫟不論在株數,頻度,密度上均占最大,其次則為長尾尖錐栗,豬腳楠,竹葉木薑仔,黃杞等:大致與喬木層之優勢種與次擾勢種相同,至其他稚樹幼昔均為本島闊葉樹林常見之植物,屬於陽性之稚樹幼苗已行絕跡,此顯示大元山常綠澗葉樹生態之演進已逐漸趨於極盛植物社會,此等值物將不失為該區之主要植物。
(5)生態演進之推論(Plant Succession)
依上述勸究結果,大元山常綠闊葉樹林生態之演進,吾人可作如是之推論,即大元山常綠闊葉樹林早期仍受人為破壞如伐採,火災,開墾等因子之影響,雖不若亞熱帶降雨林為害之烈,但林相巳遭受局部破壞,目前森林乃砍伐後發生之幼齡林,植被正在演變中,逐漸趨於極盛點(Climax),陽性植物社會早巳淘汰,現正由中性植物社會而進入較耐蔭植物社會中,即現有(1)香楠─錐果櫟群叢(2)長尾尖錐栗─豬腳楠群叢(3)南投黃肉楠─烏心石群叢(4)長尾尖錐栗一黃杞群叢等將逐漸發育旺盛,再查考其林下稚苗發生情況,仍為此四個群叢之主要植物,在不受人為干涉情況下,此四個群叢將維持大元山常綠闊葉樹林一較長久之時期,即現有森林非達安定點(Stabilization),不過為氣候控制下之極盛植物社會(Climax)發育過程中停留時期,反作用(Reaction)仍將繼續,在不受人為干涉狀態下發育大為旺盛,以改善立地環境,以適應更耐陰樹種之入侵。茲列 一簡表以示其演變順序:

3.常綠針闊葉混交林群系(Coniferous-evengreon broad leaved trees mixed forestformation)
大元山常綠針闊葉混交林自海拔1,300~2,000m,但實際上針葉樹在海拔高800m巳有發現,不過密度與頻度均小,故此林帶下部為闊葉樹林帶之延長,上部為針闊葉混交林帶,亦可稱為針葉樹與闊葉樹兩種不同植物社會之推移帶(Ecotone),針葉樹如紅檜臺灣扁柏不僅在密度大,且構成第一層樹冠為絕對優勢種,反之闊葉樹則為第二層樹冠成為被壓木,居於劣勢種地位。茲分述如下:
(1)針闊葉樹分佈概況:
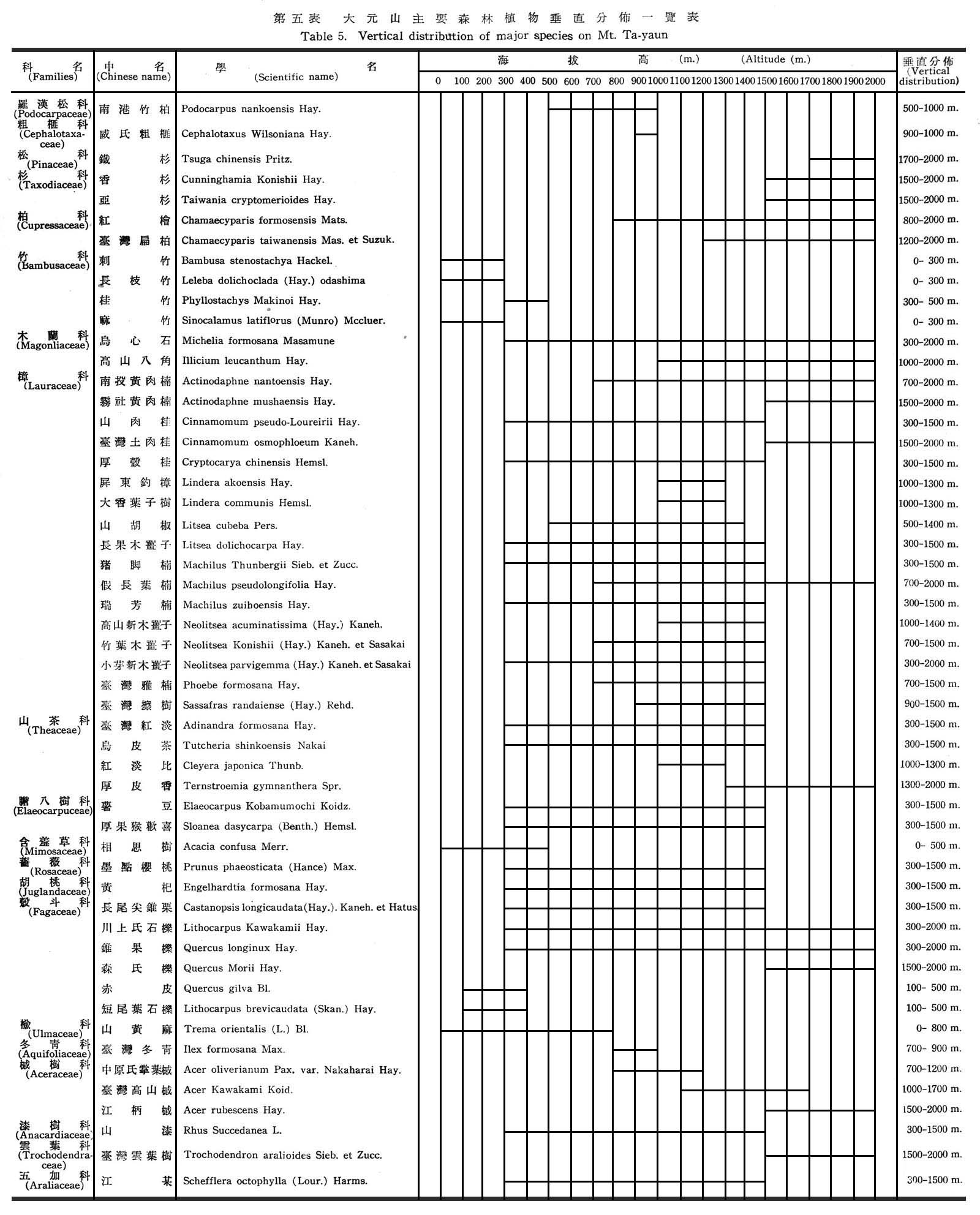
植物分佈受溫度影響者大,一般闊葉樹所需溫度較高,且生育季節亦較長,因高溫對闊葉樹馴化較低溫為易,一年中有充足時問逐漸完成其生活週期(Life Cycle),供其發生新樹冠與葉簇,反之針葉樹所需溫度低,適宣於生育季節甚短之地區,因該地區無充足時問供其發生一新樹冠與葉簇,故多在高海拔與寒冷地區發現,故針闊葉混交林帶可代表該地區針葉樹與闊葉樹所需溫度兩者之極限,以大元山論紅檜雖在850m巳有發現,但究屬少數,而闊葉樹仍占優勢,海拔高1,300~2,000m臺灣扁柏,紅檜、鐵衫、香衫、亞杉等則為真正分佈地帶,有時幾成塊狀純林,而闊葉樹林反居劣勢。茲根據大元山分場不同海拔高之各林班每木調查資料統計結果如下表所示:

由上表顯示扁柏隨海拔高逐漸增高,紅檜則以1,500~1,800m處較多,潤葉樹則海拔愈低,分佈愈盛,鐵杉分佈僅限於1,800~2,000m左右。
(2)植物群叢概述(Simple description of Plant Association)
香杉群叢(Cunninghamia Konishii Association)
分佈於海拔高1,500m,早期可能有大面積之純林,或與紅檜扁柏成混交林,目前殆已伐儘,僅於伐採跡地保留有小面積之母樹林,或偶見與紅檜扁柏,鐵杉成混交之散生林,林下多玉山箭竹,菅草、懸鉤子(Rubus Spp.)等。
鐵杉群叢(Tsuga chinensis Association)
分佈於悔拔l,800~2,000m,接近於稜線尤多,有時成純林,但多與紅檜扁柏混交,林下植物為玉山箭竹,菅草,羊齒植物等,其第二層樹則為阿里山楠(Machilus arisanensis Hay.),高山八角(Illicium leucanthum Hay.),南投黃肉楠(Actinodaphne nantoensis Hay.),錐果櫟(Quercus longinux Hay.),森氏櫟(Quercus Morii Hay.)。
紅檜及扁柏群叢(Chamaecymris Spp. Association)
分佈於海拔高1,300~2,000m,為本林帶最主要之植物社會,以海拔高1,300~1,800m處最為旺盛,其分佈最低可下降至海拔高850m之闊葉樹林中常見有老木存在,有時成純林,有時為第一層樹冠之優勢種,其第二層樹冠則為樟柯─殼斗科群叢(Lauraceae-Fagaceae Association)即所謂照葉喬木林(Laurilignosa),林下植物多玉山箭竹,菅草、羊齒植物以緻脈鳳尾草足蕨(Plagiogyria euphlebia Association)群叢占最多,溪傍及崩壞地多見有小面積純林。根據大元山分場之統計及分佈現狀之觀察,扁柏在密度及數量上較紅檜多,故生產木材亦多。
樟科―殼斗科繁叢(Lauraceae-Fagaceae Association)
分佈於海拔高1,300~2,000m,以1,300m~l,7oom處最繁殖,為構成紅檜及扁柏林之第二層樹冠,主要植物為南投黃肉楠,錐果櫟,森氏櫟,長尾尖錐栗,山肉桂、阿里山楠、豬腳楠等所組成。
亞杉群叢(Taiwania Cryptomerioides Asssociation)
臆測早期有大面積分佈,此植物乃地質時代所遺留者,目前在大元為稀有植物,僅於翠峰梅拔高1,300處偶見一二株作為母樹者。
裡白瘤足蕨單叢(Plagiagyria euplilebia, Consociation)
分佈於海拔高1,300~2,000m,為針葉樹林主要地被植物群叢之一,此群叢大體上為陰性羊齒型,主林木伐採後多自行消減,繼起者則為懸鉤子(RubusSpp.)。
懸鉤子─菅草中途群叢(Rubus Spp.-Miscanthus sinensis, Associes)
分佈于海拔高1,300~2,000m,紅檜扁柏林伐採後,首先發生者為腺萼懸鉤子(Rubes glandufoso-calycinus Hay.),苦懸鉤子(Rubus indcesuaTh•)等常與菅草混合成塊狀分佈。
冷水花―冷清草群叢(Pellionia-Pilea Spp., Association)
分佈於梅拔高1,300~1,500m,為針葉樹林林下地被植物,於濕潤環境中每多發現,主要為冷水花(Pellionia Scabra Benth.),三萼泠水花(Pellionia trilobulata Hay.),阿里山冷清草(Pilea brevicornuta Hay.)等混合組,成成塊狀分佈。
芒箕骨―石松中途群叢(Dicroanopteris dichotoma-Lycopodium spp., Associes)
分佈于海拔1,500~1,700m,土壤乾燥向陽環境中,林緣,路傍每多發現。
赤楊中途單叢(Alnus formosana, Consocies)
分佈於海拔1300~2000m,崩壞地,傾斜地,溪傍,河床地每多發生,往往成大面積帶狀分佈。
擦樹一山胡椒中途群叢(Sassafras Randaiense-Litsea Cubeba, Associes)
擦樹―懸鉤子中途群叢(Sassafras-Rubus Spp., Associes)
分佈熊梅拔高1300~1500m,土壤略乾燥之開墾地,崩壞地,開墾地每多成小團集狀態。
菅草中途群叢(Miscanthus Spp. Associes)
分佈于海拔高1300~1500m,土壤略濕潤,溪畔、溪谷、河床地等環境中適宜於生長,成小面積分佈。
玉山箭竹單叢(Pleioblastus niitakayamensis, Consociation)
分佈於海拔1300~2000m,針葉樹林林下普偏大面積分佈,或針葉樹林伐採後即行侵入,雖生長稍差但其密度益增加,利用價值少,為造林撫育上之大害。
闊葉赤車使者一達令氏雙囊蕨群叢(Elatostema edule-Athyrium Doederleinii Association)
分佈於海拔高1,300~1,500m,溪傍濕潤土壤而蔽蔭環境中,為針闊葉混交林林下常見之地被物。此群叢屬于陰性者,良好鬱閉之常綠闊葉樹林之林下,更有大面積分佈。
苔蘚植物群叢(Moss-Pogantum Spp Association)
著生於岩壁空隙處,耐乾旱亦適於濕潤,此為乾性植生列(Xerosere)之開始,依抱子而傳播,根部可分越酸性以分解岩石使產生少量之土壤,以適應雜草之侵入。
(3)組成及層次(Structure layers)
大元山針闊葉混交林之組成分子及層次,由16個樣區測定結果如下列諸表所示:
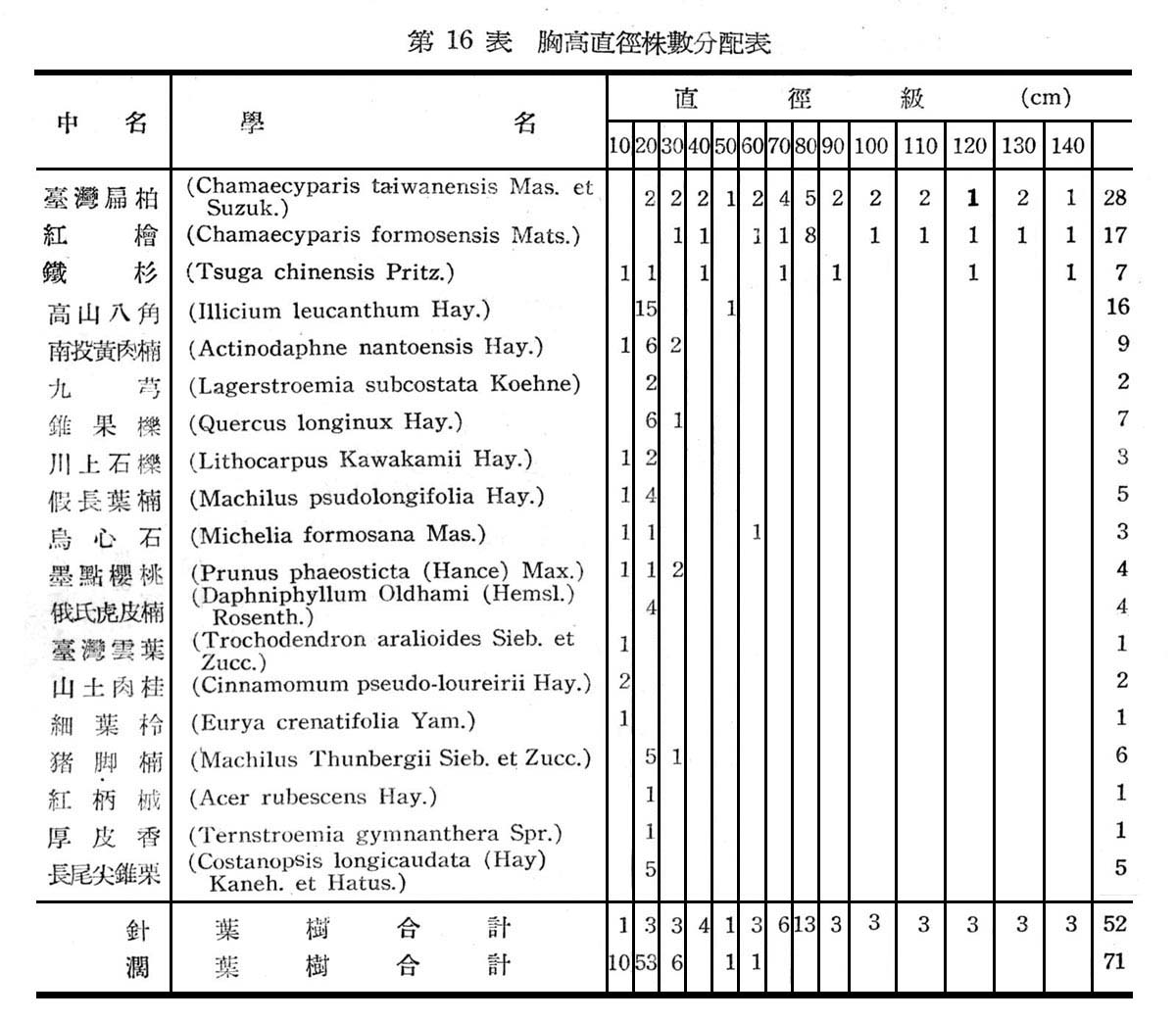
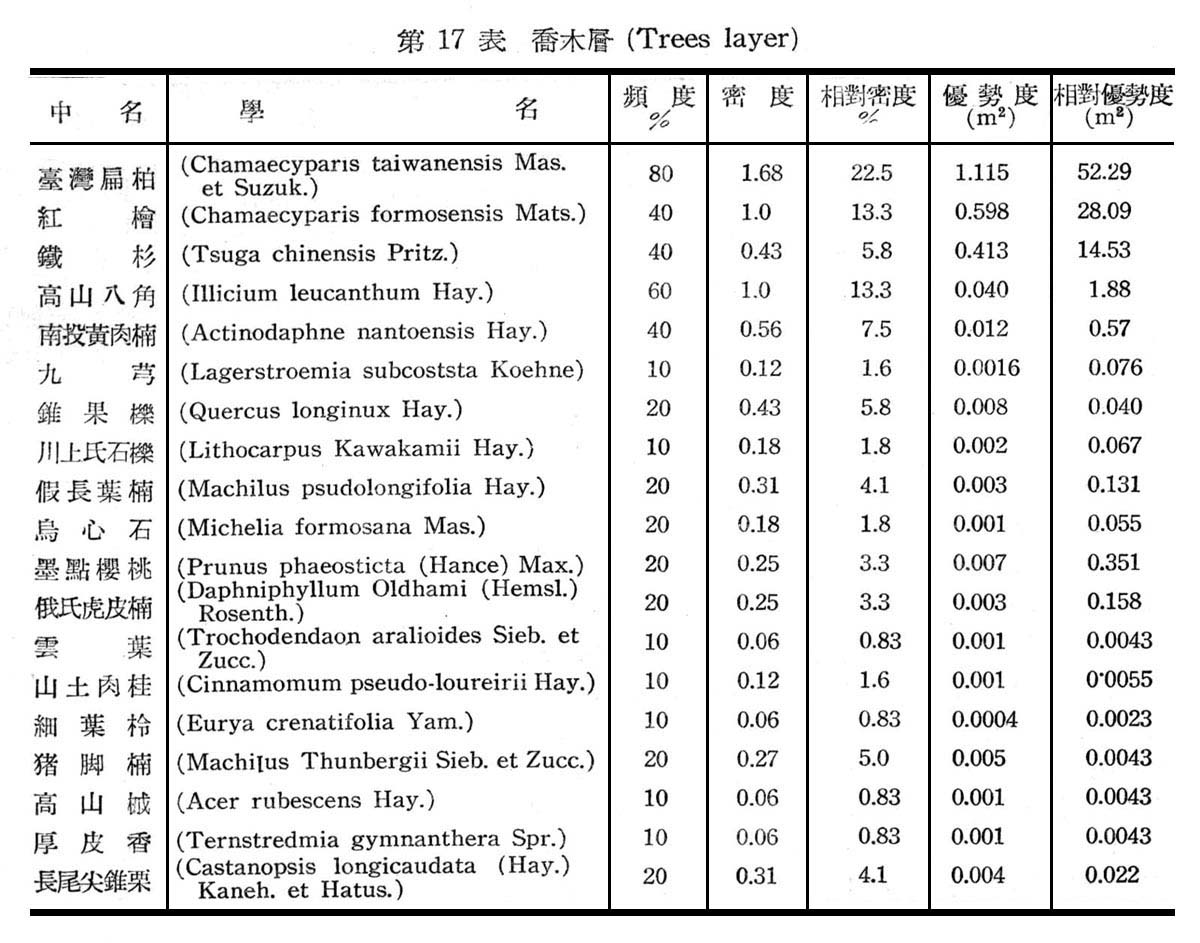

由表17一19構成大元山針闊葉混交林組成層次,計喬木層19種,灌木層18種,草木12種,羊齒植物17種,苔蘚植物4種,地衣1種。紅檜扁柏及鐵杉占絕對優勢,闊葉樹中高山八角在密度及頻度上雖大,但胸高斷面積甚小,故優勢度甚底,顯示闊葉乃後來侵入種成被壓狀態。
(4)胸高直徑與株數分配。
根據16樣區測定結果其在目前天然狀熊下,胸高直徑與株數分配如圖所示:
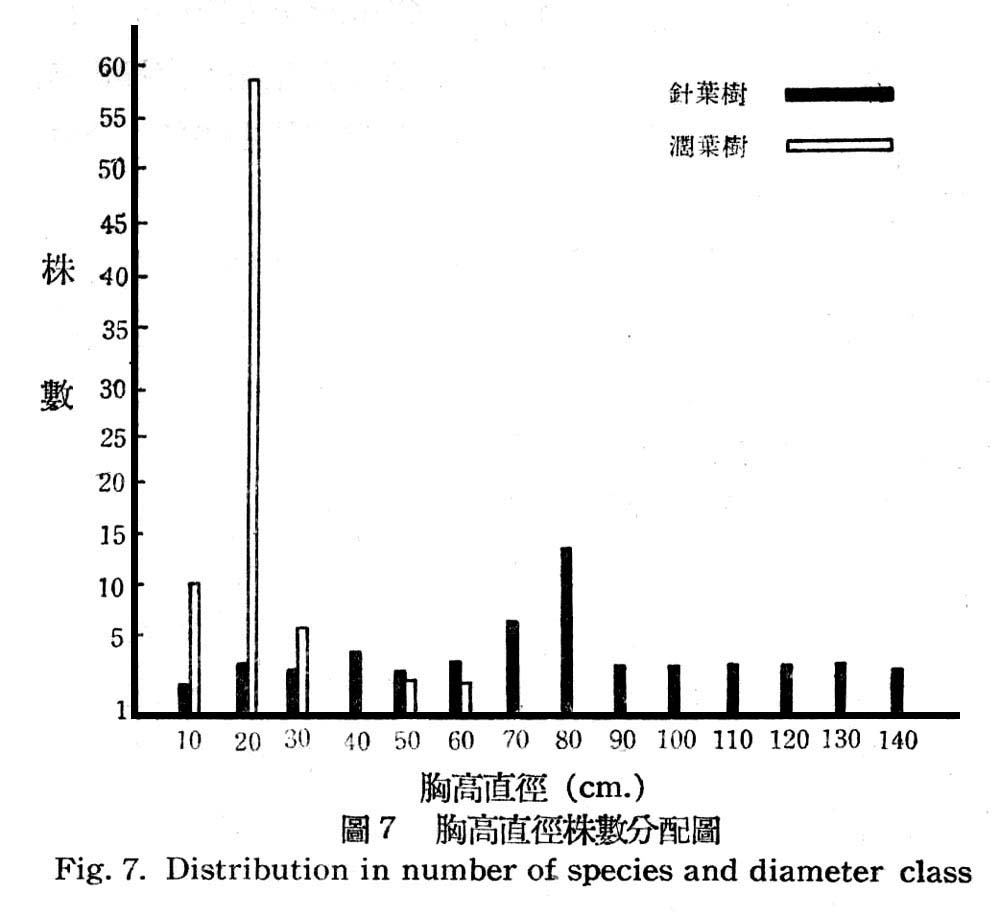
由圖7顯示闊葉樹幾全部為幼齡林,胸高直徑15~20cm占75%以上,極少數為壯齡林,反之針葉樹之林型為一齊林,即各直徑級分配頗為平均,即幼齡林,壯齡林,老齡林,三者均有平均分配,壯齡林稍多,故此早期尚未受破壞,就樹高言針葉樹平均高度均在20m以上,而闊葉樹平均高度均在15cm以下,顯示闊葉樹侵入後成壓被大。
(5)天然更新概況(Natural regeneration)
16個樣區測定其林下稚樹幼苗發生情況,給果如下表所示:
觀上表紅檜,臺灣扁柏,鐵杉在株數,密度,頻度均甚小,而稚樹幼苗發生多於崩壞地,路傍,溪畔,向陽地,枯倒木上,而在母樹林下反而少見,推究其原因,可能紅檜,臺灣扁柏,鐵杉等幼昔時期需要適當之日照,鬱閉良好之母樹下日照不足,又下層為玉山箭竹所密佈,種子難以直接著土,故稚樹幼昔均無法生長,是以在天然狀況下,針葉樹天然更新情況不佳,目前紅檜扁柏林雖然為氣候支配下之極盛植物社會(climatic Climax),恐將自行稍誠,至於闊葉樹林為針葉樹林鬱閉破壞後之侵入種,其中如高山八角,錐果櫟,南投黃肉楠等密度稍大,巳逐漸建立,目前雖為被壓木,生長不佳,如針葉樹砍伐後則將迅速占優勢,是以大元山紅檜扁柏林在目前狀況下,恐難以保持久遠,終將為闊葉樹林所取代。
(6)生態演進之推論(Plant Succession)
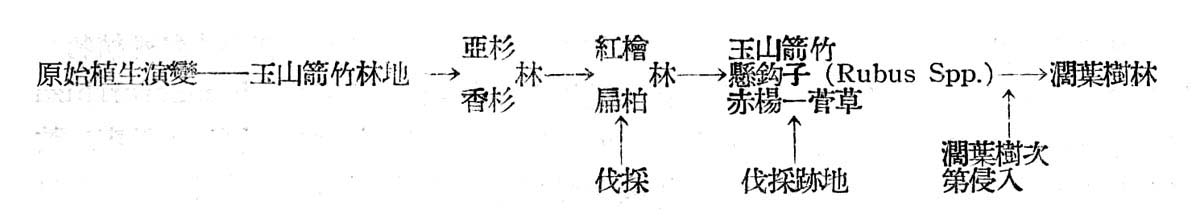
探討大元山針葉樹林之起源,並非易事,臆測紅檜扁柏林發生必較闊葉樹早,然紅檜扁柏決非古時留傳迄今者,追溯古時之植物,不易搜集,故只有注意現代植物,容視其究自古代何種典型(type)留傳於今者,若針葉樹尚不失為古時典型種,此種植物之分佈情形可賽參老,臺灣之高山早期可能為玉山箭竹所占據,以後則有亞杉與香杉所侵入,此兩種植物均為古時地質時代遺留於今者,在大元山尚有少數分佈,香杉於紅檜扁柏林伐抖勁地保留有少數,或散生於紅檜扁柏林中,亞杉則更少見僅有一二株保留作為母樹林,故紅檜扁柏林或係由亞杉香杉林演進而來,由於亞杉及香杉不耐陽光照射,在天然狀態下更新復見困難,一旦林地部分遭受破壞,因而林相有空隙處,日照射入,遂變為較亞衫及香杉耐陽光之扁柏林,然扁柏奪紅檜耐陽性差,復轉變為較扁柏尤能耐陽之臺灣紅檜林之趨勢。至于闊葉樹乃因紅檜扁柏林鬱閉破壞後之林隙部份之侵入種,因而構成扁柏紅檜林極明顯之第二層樹冠,因受紅檜扁柏樹冠之抑制,故生長及品質均不佳,在不受人為干涉天然狀態下,現時成長之紅檜扁柏,乃氣候支配下之氣候極盛植物社會(Climatic Climax),然不能保持久遠,縱使現在森林,不施伐採終其天年,林相亦將有變遷,由於紅檜扁柏天然更新之不易,在現行砍伐細度下,首先侵入者將為王山箭竹,赤楊,懸鉤子(Rubus Spp.)菅草(Miscanthus Spp.)等,再後則為上述闊葉林次第侵入,競爭與反作用結果,將變為闊葉樹林,而臺灣扁柏與紅檜將不復存在,極少可保持現狀者。
茲列表以示其演變顯序如下: